
コンテンツ
- 原核生物の遺伝情報
- 遺伝子発現の段階
- 細菌細胞の転写
- 転写:開始段階
- 転写:伸長段階
- 転写:終了フェーズ
- 細菌細胞での翻訳
- 翻訳:イニシエーション
- 翻訳:伸び
- 翻訳:終了
- 翻訳と抗生物質
- 翻訳後タンパク質処理
- タンパク質のリン酸化
- タンパク質のアセチル化とグリコシル化
- 古細菌の遺伝子発現
原核生物は、小さな単細胞生物です。これらは、2つの一般的なセルタイプの1つです。 原核生物 そして 真核生物.
原核細胞には核や細胞小器官がないため、遺伝子発現は開いた細胞質で起こり、すべての段階が同時に起こります。原核生物は真核生物よりも単純ですが、遺伝子発現の制御は細胞の挙動にとって依然として重要です。
原核生物の遺伝情報
原核生物の2つのドメインは、細菌と古細菌です。両方とも定義された核を欠いていますが、まだ遺伝暗号と核酸を持っています。真核細胞に見られるような複雑な染色体はありませんが、原核生物には核様体にデオキシリボ核酸(DNA)の円形片があります。
しかし、遺伝物質の周りに膜はありません。一般に、原核生物は、真核生物と比較して、DNAの非コード配列が少ない。これは、原核細胞が小さく、DNA分子のためのスペースが少ないためである可能性があります。
の 核様体 単に原核細胞の中でDNAが住んでいる地域です。不規則な形状をしており、サイズはさまざまです。さらに、核様体は細胞膜に付着しています。
原核生物はまた、と呼ばれる環状DNAを持つことができます プラスミド。細胞内に1つまたは複数のプラスミドを持つことが可能です。細胞分裂中に、原核生物はDNA合成とプラスミドの分離を行うことができます。
真核生物の染色体と比較して、プラスミドは小さく、DNAが少ない傾向があります。さらに、プラスミドは他の細胞DNAなしで独自に複製できます。いくつかのプラスミドは、細菌に抗生物質耐性を与えるような非必須遺伝子のコードを持っています。
特定の場合、プラスミドは、ある細胞から別の細胞に移動し、抗生物質耐性などの情報を共有することもできます。
遺伝子発現の段階
遺伝子発現とは、細胞が遺伝子コードをタンパク質生産のためにアミノ酸に翻訳するプロセスです。真核生物とは異なり、転写と翻訳の2つの主要な段階は、原核生物で同時に発生する可能性があります。
転写中、細胞はDNAをメッセンジャーRNA(mRNA)分子に変換します。翻訳中、細胞はmRNAからアミノ酸を作ります。アミノ酸がタンパク質を構成します。
転写と翻訳の両方が原核生物で起こる 細胞質。両方のプロセスを同時に行うことにより、細胞は同じDNAテンプレートから大量のタンパク質を作成できます。細胞がタンパク質をもはや必要としない場合、転写が停止する可能性があります。
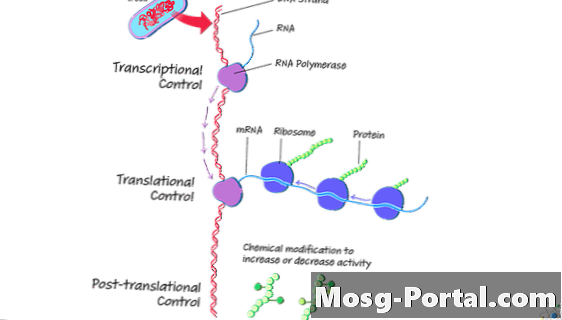
細菌細胞の転写
転写の目的は、DNAテンプレートから相補的なリボ核酸(RNA)鎖を作成することです。プロセスには3つの部分があります。開始、チェーンの伸長、終了です。
開始フェーズが発生するためには、DNAが最初に巻き戻される必要があり、これが発生する領域は 転写バブル.
細菌では、すべての転写に関与する同じRNAポリメラーゼが見つかります。この酵素には4つのサブユニットがあります。真核生物とは異なり、原核生物には転写因子がありません。
転写:開始段階
DNAがほどけてRNAポリメラーゼがプロモーターに結合すると、転写が始まります。プロモーターは、特定の遺伝子の最初に存在する特別なDNA配列です。
細菌では、プロモーターには2つの配列があります。 -10 そして -35要素。 -10エレメントは、DNAが通常ほどける場所であり、開始部位から10ヌクレオチドに位置しています。 -35要素は、サイトからの35ヌクレオチドです。
RNAポリメラーゼは、RNA転写産物と呼ばれる新しいRNA鎖を構築する際に、テンプレートとなる1つのDNA鎖に依存します。結果として生じるRNA鎖または一次転写物は、非テンプレートまたはコードDNA鎖とほぼ同じです。唯一の違いは、すべてのチミン(T)塩基がRNAのウラシル(U)塩基であることです。
転写:伸長段階
転写の鎖伸長段階で、RNAポリメラーゼはDNAテンプレート鎖に沿って動き、mRNA分子を作ります。 RNA鎖は、ヌクレオチドが追加されると長くなります。
基本的に、RNAポリメラーゼはDNAスタンドに沿って3〜5の方向に歩き、これを達成します。細菌が作り出すことができることに注意することは重要です ポリシストロン性mRNA 複数のタンパク質のコード。
•••科学転写:終了フェーズ
トランスクリプションの終了フェーズの間、プロセスは停止します。原核生物には、Rho依存性の終了とRho非依存性の終了という2つのタイプの終了段階があります。
に ローに依存する終端、Rhoと呼ばれる特別なタンパク質因子が転写を中断して終了させます。 Rhoタンパク質因子は、特定の結合部位でRNA鎖に結合します。次に、鎖に沿って移動し、転写バブル内のRNAポリメラーゼに到達します。
次に、Rhoは新しいRNA鎖とDNAテンプレートを引き離し、転写が終了します。 RNAポリメラーゼは、転写停止点であるコード配列に到達するため、移動を停止します。
に ローに依存しない終端、RNA分子はループを形成して分離します。 RNAポリメラーゼは、ターミネーターであり、多くのシトシン(C)およびグアニン(G)ヌクレオチドを持つテンプレート鎖上のDNA配列に到達します。新しいRNA鎖は、ヘアピン型に折り畳まれ始めます。そのCおよびGヌクレオチドが結合します。このプロセスにより、RNAポリメラーゼの移動が停止します。
細菌細胞での翻訳
翻訳は、転写中に作成されたRNAテンプレートに基づいてタンパク質分子またはポリペプチドを作成します。細菌では、翻訳はすぐに起こることがあり、時には転写中に開始されます。これは、原核生物にはプロセスを分離するための核膜やオルガネラがないためです。
真核生物では、転写は核で起こり、翻訳は サイトゾル、または細胞の細胞内液。真核生物はまた、翻訳前に処理される成熟mRNAを使用します。
バクテリアで翻訳と転写が同時に起こりうる別の理由は、真核生物で見られる特別なプロセシングをRNAが必要としないことです。細菌RNAはすぐに翻訳の準備ができています。
mRNAの鎖はと呼ばれるヌクレオチドのグループを持っています コドン。各コドンには3つのヌクレオチドがあり、特定のアミノ酸配列をコードします。アミノ酸は20個しかありませんが、細胞には61個のコドンと3つの停止コドンがあります。 AUGは開始コドンであり、翻訳を開始します。また、アミノ酸メチオニンをコードします。
翻訳:イニシエーション
翻訳中、mRNA鎖はタンパク質になるアミノ酸を作るためのテンプレートとして機能します。細胞はこれを達成するためにmRNAを解読します。
開始には、トランスファーRNA(tRNA)、リボソーム、およびmRNAが必要です。各tRNA分子には アンチコドン アミノ酸用。アンチコドンはコドンに相補的です。細菌では、小さなリボソーム単位がmRNAに結合するとプロセスが始まります。 Shine-Dalgarnoシーケンス.
Shine-Dalgarno配列は、細菌と古細菌の両方における特別なリボソーム結合領域です。あなたは通常、開始コドンAUGから約8ヌクレオチドを見ます。
細菌の遺伝子はグループで転写を起こすことができるため、1つのmRNAが多くの遺伝子をコードする場合があります。 Shine-Dalgarnoシーケンスにより、開始コドンを見つけやすくなります。
翻訳:伸び
伸長中、アミノ酸の鎖は長くなります。 tRNAはアミノ酸を付加してポリペプチド鎖を作ります。 tRNAは Pサイト、これはリボソームの中間部分です。
Pサイトの横には サイト。コドンに一致するtRNAは、Aサイトに移動できます。次に、アミノ酸の間にペプチド結合が形成されます。リボソームはmRNAに沿って動き、アミノ酸は鎖を形成します。
翻訳:終了
停止コドンが原因で終了します。停止コドンがAサイトに入ると、停止コドンには相補的なtRNAがないため、翻訳プロセスが停止します。呼ばれるタンパク質 リリース要因 P部位に適合すると、停止コドンが認識され、ペプチド結合の形成が防止されます。
これは、放出因子が酵素に水分子を追加させ、tRNAから鎖を分離させるために起こります。
翻訳と抗生物質
感染症を治療するために抗生物質を摂取すると、細菌の翻訳プロセスを妨害して機能する場合があります。抗生物質の目的は、バクテリアを殺し、バクテリアの繁殖を防ぐことです。
彼らがこれを達成する一つの方法は、細菌細胞のリボソームに影響を与えることです。薬物はmRNAの翻訳を妨害したり、ペプチド結合を作る細胞の能力をブロックしたりする可能性があります。抗生物質はリボソームに結合できます。
たとえば、テトラサイクリンと呼ばれる抗生物質の一種は、原形質膜を通過して細胞質内に蓄積することにより、細菌細胞に入ることができます。次に、抗生物質はリボソームに結合して翻訳をブロックします。
シプロフロキサシンと呼ばれる別の抗生物質は、複製を可能にするためにDNAをほどく役割を担う酵素を標的とすることにより、細菌細胞に影響を与えます。どちらの場合でも、人間の細胞は節約され、人々は自分の細胞を殺さずに抗生物質を使用できます。
関連トピック: 多細胞生物
翻訳後タンパク質処理
翻訳が終了した後、一部の細胞はタンパク質の処理を続けます。 翻訳後修飾 タンパク質の(PTM)により、細菌は環境に適応し、細胞の挙動を制御できます。
一般に、原核生物ではPTMは真核生物ほど一般的ではありませんが、一部の生物にはPTMがあります。細菌はタンパク質を修飾し、プロセスを逆転させることもできます。これにより、より汎用性が増し、調節のためにタンパク質修飾を使用できるようになります。
タンパク質のリン酸化
タンパク質リン酸化 細菌の一般的な修飾です。このプロセスでは、リン原子と酸素原子を含むリン酸基をタンパク質に追加します。リン酸化はタンパク質の機能に不可欠です。
ただし、リン酸化は可逆的であるため、一時的な場合があります。一部の細菌は、他の生物に感染するプロセスの一部としてリン酸化を使用できます。
セリン、スレオニン、チロシンのアミノ酸側鎖で起こるリン酸化は Ser / Thr / Tyrリン酸化.
タンパク質のアセチル化とグリコシル化
リン酸化タンパク質に加えて、細菌は アセチル化 そして グリコシル化 タンパク質。また、メチル化、カルボキシル化、およびその他の修飾が行われる場合があります。これらの修飾は、細菌の細胞シグナル伝達、調節、およびその他のプロセスで重要な役割を果たします。
たとえば、Ser / Thr / Tyrリン酸化は、細菌が環境の変化に反応し、生存の可能性を高めるのに役立ちます。
研究によると、細胞の代謝変化はSer / Thr / Tyrリン酸化に関連していることが示されており、これは細菌が細胞プロセスを変化させることで環境に応答できることを示しています。さらに、翻訳後の変更により、迅速かつ効率的に対応できます。変更を元に戻す機能も重要な制御を提供します。
古細菌の遺伝子発現
古細菌は、真核生物により類似した遺伝子発現メカニズムを使用します。古細菌は原核生物ですが、遺伝子発現や遺伝子調節など、真核生物と共通するものがいくつかあります。古細菌の転写と翻訳のプロセスも、細菌といくつかの類似点があります。
例えば、古細菌と細菌の両方は、最初のアミノ酸としてメチオニンを、開始コドンとしてAUGを持っています。一方、古細菌と真核生物の両方には TATAボックス、これはプロモーター領域のDNA配列で、DNAをデコードする場所を示します。
古細菌の翻訳は、細菌に見られるプロセスに似ています。どちらのタイプの生物も、30Sおよび50Sサブユニットという2つのユニットで構成されるリボソームを持っています。さらに、両方ともポリシストロン性mRNAとシャインダルガルノ配列を持っています。
細菌、古細菌、真核生物には複数の類似点と相違点があります。しかし、それらはすべて、生き残るために遺伝子発現と遺伝子調節に依存しています。