コンテンツ
生物学の初心者でも長年の愛好家でも、デフォルトでは、デオキシリボ核酸(DNA)をライフサイエンスのすべてで最も不可欠な概念と見なす可能性があります。少なくとも、DNAは地球上の何十億という人々の中であなたをユニークにするものであり、分子生物学の講義の中心的なステージであると同時に刑事司法の世界でも役割を果たしていることをご存知でしょう。あなたは、DNAが両親から受け継いだあらゆる特性をあなたに与える責任があり、あなた自身のDNAが子供がいれば将来の世代への直接の遺産であることをほぼ確実に学びました。
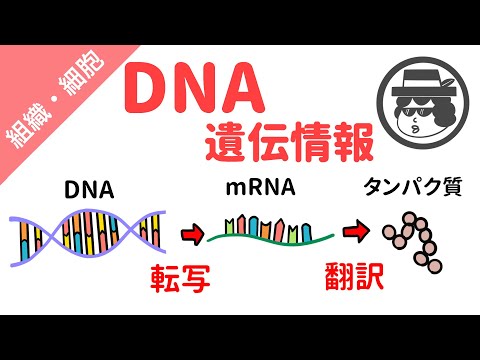
あなたがあまり知らないかもしれないのは、細胞内のDNAを、顕在化と隠蔽の両方で明らかにする身体的特徴に結びつける経路と、その経路に沿った一連のステップです。分子生物学者は、その分野で「中央教義」という概念を生み出しました。これは単に「DNA to RNA to protein」と要約できます。このプロセスの最初の部分-DNAからRNAまたはリボ核酸を生成-は、 転写、そしてよく研究され調整されたこの一連の生化学体操は、科学的に深遠であると同時にエレガントです。
核酸の概要
DNAとRNAは核酸です。どちらも人生の基本です。これらの高分子は非常に密接に関連していますが、それらの機能は非常に密接に絡み合っていますが、非常に多様で特殊です。
DNAはポリマーであるため、多数の繰り返しサブユニットで構成されています。これらのサブユニットは正確に同一ではありませんが、形式は同一です。 4色の立方体で構成されたビーズの長いストリングを考えてみましょう。サイズは非常にわずかに異なり、DNAとRNAがどのように配置されているかについての基本的な感覚が得られます。
核酸のモノマー(サブユニット)は ヌクレオチド。ヌクレオチド自体は、3つの異なる分子のトライアドで構成されます:リン酸基(または複数のグループ)、5炭素糖、窒素に富む塩基(「基礎」の意味ではなく「水素イオン受容体」を意味する「塩基」) )。核酸を構成するヌクレオチドには1つのリン酸基がありますが、2つまたは3つのリン酸が連続して結合しているものもあります。分子アデノシン二リン酸(ADP)とアデノシン三リン酸(ATP)は、細胞エネルギー代謝において非常に重要なヌクレオチドです。
DNAとRNAはいくつかの重要な点で異なります。 1つは、これらの各分子には4つの異なる窒素塩基が含まれ、DNAにはアデニン(A)、シトシン(C)、グアニン(G)、チミン(T)が含まれ、RNAにはこれらの最初の3つが含まれますが、ウラシル(U) T. 2の場合、DNAの糖はデオキシリボースであり、RNAの糖はリボースです。そして3つ目は、DNAが最もエネルギー的に安定した形で二本鎖であるのに対し、RNAは一本鎖です。これらの違いは、転写および特異的にこれらの各核酸の機能の両方において一般的に非常に重要です。
塩基AおよびGはプリンと呼ばれ、C、TおよびUはピリミジンとして分類されます。重大なことに、AはT(DNAの場合)またはU(RNAの場合)にのみ化学的に結合します。 CはGにのみ結合します。DNA分子の2本の鎖は相補的です。つまり、各鎖の塩基はすべての点で反対の鎖のユニークな「パートナー」塩基と一致します。したがって、AACTGCGTATGはTTGACGCATAC(またはUUGACGCAUAC)を補完します。
DNA転写と翻訳
DNA転写のメカニズムを掘り下げる前に、DNAとRNAに関連する用語を確認するために時間を割く価値があります。ミックス内の非常に類似した発音の単語が多いため、それらを混同しやすいからです。
複製 何かの同一のコピーを作成する行為です。書面の文書のコピーを作成する(古い学校)か、コンピューターでコピーアンドペースト機能を使用する(新しい学校)と、両方の場合にコンテンツを複製します。
DNAは複製されますが、RNAは、現代の科学が確認できる限り、しません。書き起こしからのみ発生します。_「ラテン語の書き方」を意味するラテン語のルートから、書き起こしは元のソースのコピーの特定のエンコードです。音声記録として作成された医療メモを書面に入力することが仕事である医療記録転写士のことを聞いたことがあるかもしれません。理想的には、言葉、したがってが、媒体が変わってもまったく同じになります。細胞では、転写は窒素塩基配列の言語で書かれた遺伝子DNAのRNA形式、特にメッセンジャーRNA(mRNA)へのコピーを伴います。このRNA合成は真核細胞の核で起こり、その後、mRNAは核を離れ、リボソームと呼ばれる構造に向かいます。 翻訳.
転写は異なる媒体での単純な物理的エンコードであるのに対し、生物学的用語での翻訳は、それを意図的なアクションに変換することです。と呼ばれるDNAまたは単一のDNAの長さ 遺伝子、最終的に細胞はユニークなタンパク質製品を製造します。 DNAはこれをmRNAの形で出荷します。その後、mRNAはリボソームに運ばれ、タンパク質に変換されます。このビューでは、mRNAは青い家具または家具を組み立てるための一連の指示に似ています。
うまくいけば、核酸が何をするかについてあなたが持っている謎が明らかになるでしょう。しかし、特に転写はどうですか?
転写のステップ
むしろ有名なDNAは、二本鎖のらせんに織り込まれています。しかし、この形式では、そこから何かを構築することは物理的に難しいでしょう。したがって、 開始 転写の段階(またはステップ)で、DNA分子はヘリカーゼと呼ばれる酵素によってほどかれます。結果として生じる2本のDNA鎖のうち、一度に1つだけがRNA合成に使用されます。このストランドは ノンコーディング なぜなら、DNAとRNAの塩基対合の規則のおかげで、もう一方のDNA鎖は合成されるmRNAと同じ窒素塩基の配列を持っているため、この鎖が コーディング ストランド。以前に行ったポイントに基づいて、製造に関与するDNA鎖とmRNAは相補的であると結論付けることができます。
ストランドの動作準備ができたら、プロモーターシーケンスと呼ばれるDNAのセクションに、ストランドに沿って転写が開始される場所が示されます。酵素RNAポリメラーゼはこの場所に到着し、プロモーター複合体の一部になります。これはすべて、mRNA合成がDNA分子上で正確に開始されることを保証することであり、これにより目的のコード化されたを保持するRNA鎖が生成されます。
次に、 伸長 フェーズでは、RNAポリメラーゼがDNA鎖を「読み取り」、プロモーター配列から始まりDNA鎖に沿って移動します。先生が生徒の列を歩いてテストを配布し、新しく形成されるRNA分子の成長端にヌクレオチドを追加するように。
1つのヌクレオチドのリン酸基と次のヌクレオチドのリボースまたはデオキシリボース基の間に作成される結合は ホスホジエステル結合。 DNA分子には、一方の端に3(「スリープライム」)末端、もう一方の端に5(「ファイブプライム」)末端と呼ばれるものがあり、これらの数字は末端の炭素原子位置に由来することに注意してくださいそれぞれの末端リボース「リング」。 RNA分子自体が3方向に成長すると、DNA鎖に沿って5方向に移動します。 mRNA合成のメカニズムを完全に理解していることを確認するには、図を調べる必要があります。
ヌクレオチドの追加–具体的には、ヌクレオシド三リン酸(ATP、CTP、GTPおよびUTP; ATPはアデノシン三リン酸、CTPはシチジン三リン酸など)–伸長mRNA鎖へのエネルギーが必要です。これは、非常に多くの生物学的プロセスと同様に、ヌクレオシド三リン酸自体のリン酸結合によって提供されます。高エネルギーのリン酸-リン酸結合が切断されると、得られたヌクレオチド(AMP、CMP、GMP、UMP。これらのヌクレオチドでは、「MP」は「モノリン酸」を表します)がmRNAと無機リン酸分子のペアに追加されます、通常書かれたPP私、落ちます。
転写が起こると、前述のように、DNAの一本鎖に沿って転写します。ただし、DNA分子全体がほどけて相補鎖に分離することはありません。これは、転写のすぐ近くでのみ起こります。その結果、DNA分子に沿って移動する「転写バブル」を視覚化できます。これは、1つのメカニズムによってオブジェクトの直前に解凍されているジッパーに沿って移動するオブジェクトに似ていますが、別のメカニズムがオブジェクトウェイクでジッパーを再圧縮します。
最後に、mRNAが必要な長さと形に達すると、 終了 フェーズが進行中です。開始と同様に、このフェーズは、RNAポリメラーゼの停止標識として機能する特定のDNAシーケンスによって有効になります。
細菌では、これは2つの一般的な方法で発生します。これらの1つでは、終止配列が転写され、それ自体に折り返され、それによってRNAポリメラーゼがその仕事を続けているときに「束ねられる」mRNAの長さが生成される。これらのmRNAの折り畳まれた部分は、しばしばヘアピン鎖と呼ばれ、一本鎖であるがゆがめられたmRNA分子内の相補的塩基対合を含む。このヘアピンセクションの下流には、延長されたU塩基または残留物があります。これらのイベントにより、RNAポリメラーゼはヌクレオチドの追加を停止し、DNAから切り離され、転写が終了します。これは、rho因子として知られるタンパク質に依存しないため、rho非依存性終結と呼ばれます。
rho依存性終結では、状況はより単純であり、ヘアピンmRNAセグメントまたはU残基は必要ありません。代わりに、rho因子はmRNA上の必要なスポットに結合し、RNAポリメラーゼからmRNAを物理的に引き離します。 rho非依存性またはrho依存性の終了が起こるかどうかは、DNAおよびmRNA(さまざまなサブタイプが存在する)に作用しているRNAポリメラーゼの正確なバージョン、および直接の細胞環境におけるタンパク質およびその他の要因に依存します。
両方のイベントのカスケードは、最終的に転写バブルでDNAを破壊するmRNAをもたらします。
原核生物対真核生物
原核生物(ほとんどすべてが細菌)と真核生物(動物、植物、真菌などの多細胞生物)の転写には多くの違いがあります。例えば、原核生物での開始は通常、プリブノーボックスとして知られるDNA塩基配列を含み、TATAATの塩基配列は転写開始自体が起こる場所からおよそ10塩基対離れています。しかし、真核生物には、開始部位からかなり離れた位置にエンハンサー配列があり、RNAポリメラーゼによりアクセスしやすいようにDNA分子を変形させる活性化タンパク質があります。
さらに、細菌では、真核生物(毎分約22〜25塩基対)の約2倍の速さ(毎分約42〜54塩基対、1秒間に1境界)で伸長が起こります。最後に、細菌の終結のメカニズムは上記で説明されていますが、真核生物では、このフェーズには特定の終結因子、およびポリAと呼ばれるRNAの鎖(多くのアデニン塩基が「尾」と呼ばれます)が含まれます。伸長の停止がmRNAのバブルからの切断を引き起こすのか、切断自体が伸長プロセスを突然終了させるのかはまだ明らかではありません。